RESEARCH
ウイルス感染等に対する生命システムの応答機構の解明と予防医療、ワクチン・感染症薬開発への基盤的研究
近年、新型コロナウイルス (SARS-CoV2)による感染症(COVID-19)、新型肺炎(SARS)、中東呼吸器症候群 (MERS)やH5N1鳥インフルエンザウイルスによる高病原性インフルエンザウイルス感染症などの、ヒトに重篤な生命予後をもたらすことのあるウイルス感染症が、数十年おきに発生しています。また病原性の高くないウイルスによる感染でも、ヒト(宿主)が基礎疾患や加齢等のリスクファクターを有していると重症化することがあります。一旦重症化すると、集中治療室 (ICU)で人工呼吸や膜型人工肺(ECMO)などの救命治療が必要になります。このように、新型コロナやインフルエンザなどのウイルス感染症では、無症状や軽症から、人工呼吸が必要になるほどの重症、さらにECMOが必要になるほどの超重症まで多様な感染病態が形成されます。ここにはウイルスゲノムで規定されるウイルス側の因子に加え、宿主側の遺伝的・非遺伝的因子の双方が関わっています。宿主側の非遺伝的因子には加齢や環境要因等と関連したエピジェネティックな因子が含まれます。さらに最近、このような重症ウイルス感染症ではICUを退院した後も、ICUフレイル(ICU後症候群 (PICS); post ICU syndrome)とよばれるサルコペニア、うつ、認知障害を特徴とする後遺症が引き起こされ、特に高齢者においてQOL(Quality of Life)が著しく低下することが問題になっています。これまでインフルエンザウイルスやSARS-CoV2に対してウイルスタンパク質を標的とした様々な抗ウイルス薬が開発されてきました。これらは基本的にはウイルスの生活環に介入するもので、変異ウイルスでは効果が消失する可能性があり、またウイルス産生のピークを過ぎた後に宿主側で引き起こされる重症化や後遺症を直接制御するものではありません。そこでこのアプローチに加え、変異ウイルスを含むより広い範囲のウイルスに効果を示し、重症化や後遺症自体を制御可能な感染症薬が求められています。
生命システムがウイルスの侵襲を受けると、ウイルスと宿主の相互作用によって、宿主においては、分子スケールでは、遺伝的あるいは非遺伝的(エピジェネティックな)要因が関わるマルチオミクスのネットワークが、個体スケールでは生体機能の変化を介した多彩な病態が形成されます。私達は、感染系を用いた生物学的解析と臨床ビッグデータを用いたデータ駆動型解析とを双方向性に連動させて、ウイルスに対する生命体のシステムとしての応答機構について、クロマチン・エピジェネティクス制御機構に焦点を当てて研究しています。データ駆動型解析では、ウイルスに感染したヒトの診療情報、ウイルスゲノム情報、オミクス情報、バイタルデータ等を収集・生成し、これらマルチモーダルなデータの解析プラットフォームを構築しています。これを用いて、急性期の重症化ならびに長期の後遺症について、病態形成のネットワークを同定し、病態、介入効果を予測するAIモデル等を構築しています。これらを通して、個人に最適な予防法や治療法の開発といった感染症精密医療に資する研究基盤の構築を目指しています(図1)。

1. ウイルス感染に対する生命システムの応答機構の解明とエピゲノム創薬・予防医療への応用
インフルエンザウイルスは、ゲノムの転写・複製を核内で行うRNAウイルスですが、ウイルスが感染した宿主細胞の染色体にはウイルス・宿主相互作用を介した連続的な摂動 (perturbation)が加わることになります。近年、外的要因による染色体高次構造の変化とエピジェネティックな遺伝子制御の関係が注目されています。染色体は、遺伝子転写、複製、組換え、修復、分配、エピゲノム修飾などの各種機能を通じて生命活動に深く関わっていますが、DNAメチル化やヒストン化学修飾に代表されるエピジェネティックな遺伝子発現調節に加え、プロモーター、エンハンサー、サイレンサー、インスレーターなどの遺伝子制御領域の相互作用に関わる染色体高次構造が、エピジェネティックな遺伝子発現制御の重要な要素であることがわかってきました。核内には遺伝情報を含む染色体テリトリーに加え、PMLボディー、核小体、核膜等数々の構造体が存在しますが、それらが空間内でどのように形成され相互作用するかは、遺伝子の効率的な発現やエピジェネティックな制御を規定していると考えられています。私達は、ウイルス感染という外的要因による染色体高次構造の変化と、各種染色体機能の連携に興味を持っています。
ヒトの細胞は直径数ミクロンの核を有しており、そこに全長2メートルにもおよぶDNAがクロマチンを形成して収納されています。この限られた空間内で、クロマチンは不規則に折りたたまれて、ループやTAD(topologically associating domain)とよばれるドメインからなる3D構造を取っています。また最近、間期においては、ループやTAD等のクロマチン構造は、コヒーシンと呼ばれるクロマチンを束ねるリング状のタンパク質により維持されていることが分かってきました。これまでクロマチンの 3D 構造は動的に変化することが知られており、インフルエンザウイルス感染においてはウイルスが宿主の核内タンパク質やクロマチンを構成しているヌクレオソームと相互作用することが知られていました。また、H4K20のトリメチル化酵素であるSuv4-20h2はクロマチン構造の維持に関っているコヒーシンと結合していることが報告されています。しかし、インフルエンザウイルス感染にともなったSuv4-20h2やコヒーシンの細胞内動態、さらにこれらが宿主クロマチンの 3D 構造のダイナミクス、さらにインフルエンザの病態の形成にどのような影響を与えるかは不明でした。
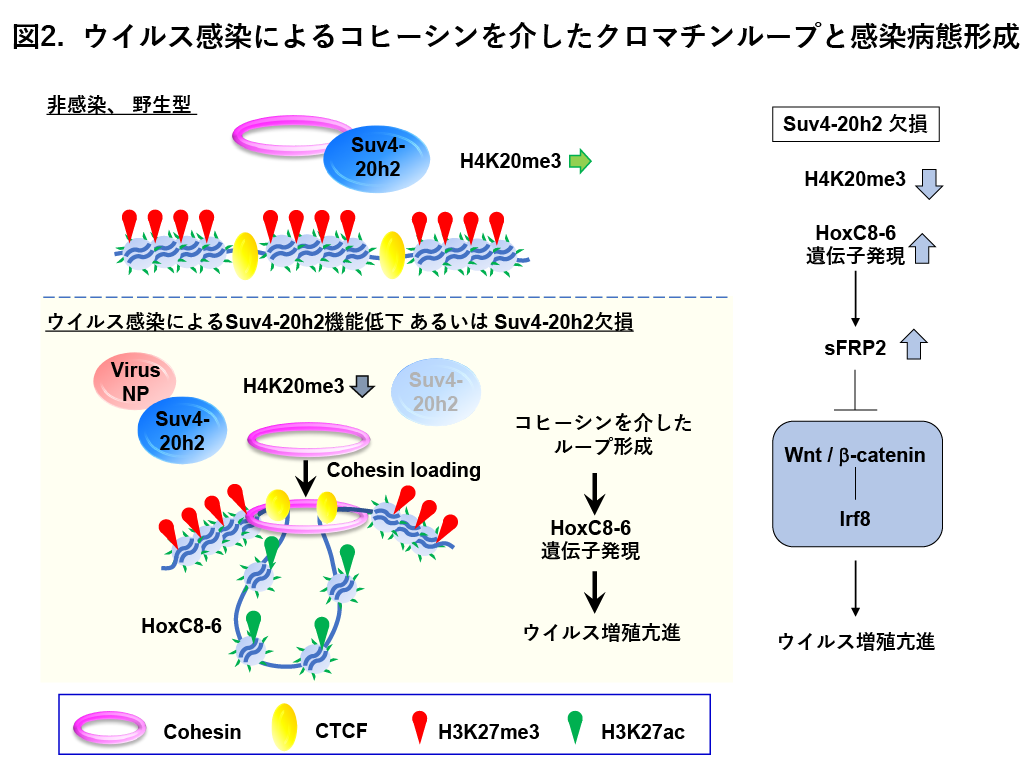
私達は、インフルエンザウイルス感染にともなって、ウイルスタンパク質とH4K20のトリメチル化酵素であるSuv4-20h2が相互作用することによって、Suv4-20h2の機能が低下してH4K20のトリメチル化(H4K20me3) レベルが低下することを発見しました。Suv4-20h2は非感染状態では、クロマチン構造の維持に関っているコヒーシンと結合していますが、感染に伴ってSuv4-20h2とコヒーシンの結合が低下して、特定のゲノム領域(HoxC8-6)のバウンダリーにコシーシンがローディングして、クロマチンループが形成され、この時同領域の遺伝発現が誘導されることがわかりました(図2)。HoxC8-6は、Wnt/β-catenin経路を阻害することで、インターフェロン制御因子であるIRF8の発現を抑制し、ウイルス増殖の亢進することを見出しました(図2)。Suv4-20h2を欠損させたインフルエンザ感染マウスでは、生存率やウイルスの増殖、肺の病理所見の悪化を認め、インフルエンザの病態が悪化することが分かりました。さらに、肺がん患者では、Suv4-20h2ならびにH4K20me3レベルが低下しており、さらに肺がん細胞ではインフルエンザウイルスの増殖が亢進することがわかりました。これらの結果から、Suv4-20h2はコヒーシンを介したクロマチンループ形成を制御してインフルエンザに保護的に作用することがわかりました。Suv4-20h2によるコヒーシンを介したクロマチンループ形成等のエピジェネティクス制御はインフルエンザの新しい治療法やバイオマーカー(H4K20me3など)の開発につながることが示唆されます。これらの研究成果は、2021年6月『iScience』に掲載されました。
2. 臨床マルチモーダルデータの生成・データ駆動型解析による重症化メカニズムの解明と予防医療・創薬応用
新型コロナウイルス感染症の臨床データシェアリングを促すために、R2年度SIPのAIH研究で、二次利用が可能なクラウド型のデータベースを構築しました。R3年度はこのデータベースのデータ提供を行いました。クラウドを用いた医療データベースの提供は、国内初のものです。また、新型コロナAIシミュレーションプロジェクトでは、ICUに入院したコロナ患者 1,794例の複数時点の診療情報をもとに、生死/重症度を予測する機械学習モデルを作成しました。長期予測モデルとしては、入院時の診療情報から、退院時の生死を高い精度で予測することができました。短期予測モデルでは、入院中のある時点での診療情報から、n日後の重症度を予測するもので、⇒例えば、5日後にECMOが必要になったり、あるいはECMOから離脱できることが高い精度で予測可能となりました。これは、重症患者の治療に加え、新型コロナの医療提供体制(ECMO・感染病床の確保など)の整備に有用であると思われます。さらに、ICUに入ったCOVID-19患者の診療情報, バイタル情報, マルチオミクスデータを生成し、これらマルチモーダルなデータの解析のパイプラインを構築しました。診療情報に加え、ICU統合型医療情報システムから血圧、心拍、パルスオキシメーター等のリアルタイムのバイタル情報を収集しています。これらを用いて、病態変化の予兆を捉えるDynamical Network Biomarker (DNB)の解析を行っています。また、PBMCを用いたシングルセルマルチオーム解析では、高齢者においては、非感染状態で既に、免疫学的多様性が高いことを報告しました。これらは、個人の多様性を重視した予防医療や感染症創薬につながるものです。
3. ウイルス感染における宿主脂溶性代謝と核内ウイルスRNAの輸送制御
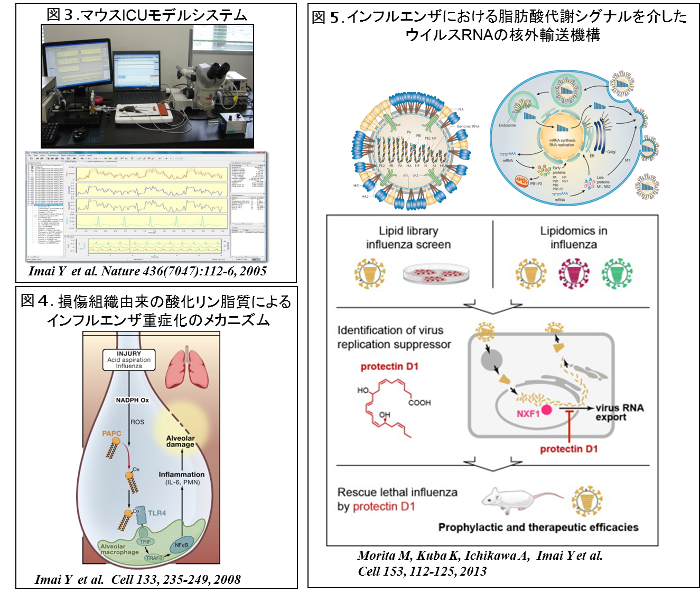
ウイルスは宿主の細胞内小器官を利用して増殖するので、ウイルスの増殖は宿主因子に依存します。ウイルスが侵入した宿主細胞においては、ウイルスと宿主の相互作用から様々なネットワークが可動し、この複雑系のネットワークによってウイルスによる生体侵襲が調節されていると考えられます。これまでウイルスによって生体が高度の侵襲を受けるメカニズム、すなわちウイルス感染症の重症化を制御している分子機構は十分に解明されていません。私達の研究室では、ウイルスに感染して重症化した患者さんがICUで集中治療を受ける状態を再現することが可能なマウスICUモデル(図3)を独自に樹立し、このモデルを用いて、新型肺炎SARSウイルスやH5N1鳥インフルエンザウイルスに感染した肺組織においては、損傷組織由来の酸化リン脂質が損傷関連分子パターン(DAMPs)として作用することによって、自然免疫が過剰に活性化され、制御範囲を逸脱した重度の組織炎症が引き起こされ、致死的な呼吸不全に至ることを報告しました (図4)(Cell 2008)。
ところで、インフルエンザウイルスと宿主の相互作用に関しては、トランスクリプトーム、プロテオームの解析が既に行われていましたが、生体内化合物はほとんど注目されていませんでした。そこで私達は、脂肪酸代謝物のライブラリーを用いてインフルエンザウイルスの増殖に関するスクリーニングを行い、増殖を抑制する新規のn-3系脂肪酸代謝物を同定しました。これは、化合物のライブラリーを用いて、インフルエンザウイルスを感染させたヒト肺上皮 A549細胞にそれぞれの化合物を投与し、ウイルスタンパク質のmRNA発現量を指標に、96ウェルフォーマットでスクリーニングを行うものです。次にスクリーニングで増殖の抑制が見られた化合物に関して、ウイルスを経気道的にマウス肺に感染させた重症インフルエンザモデルを用いて、化合物のin vivoでの有効性を確認しました。次いで、感染マウスの肺組織を用いてリピドミクス解析を行いました。さらに、同代謝物がウイルスの増殖を抑えるメカニズムの手がかりを得るために、マイクロアレイのデータを用いたIngenuity pathway analysis (IPA)を行いました。これはオミクスデータをもとにして生物学的な機能の解釈やパスウェイ解析を行うことができるソフトウェアです。この解析で同代謝物の投与によってRNA transportのパスウェイが変動しているというおもしろい結果を得ました。そこでウイルスRNAの細胞内局在をRNA-FISH法で解析したところ、同代謝物を投与すると、確かにウイルスRNAの核外輸送が抑えられることがわかりました。さらにRNAゲルシフトアッセイ、RNA免疫沈降(RIP)、RIP-seq等を通して、同代謝物はウイルスRNAがRNA核外輸送因子NXF1にリクルートされる過程を抑えることによってウイルスの増殖を抑制していることを見出しました。興味深いことに、同代謝物とノイラミニダーゼ阻害薬といった作用点の異なる2剤を併用すると、これまで救命が難しかった、感染から48時間以降に投与を開始しても、重症インフルエンザマウスの生存率を改善させることができました。これらの研究成果はCell誌に報告しました (図5)。その後、NEJMは"Influenza Time to Target the Host"として我々の仕事を紹介し、宿主を標的とした新しい抗インフルエンザ薬の可能性を指摘しています。
4. ウイルス感染の重症病態の形成を司る動的ネットワークの予測と先制治療への応用
インフルエンザウイルスは、高病原性ウイルス(例:H5N1ウイルス)によって、あるいは低病原性ウイルスであっても宿主側の要因(例:糖尿病の合併)によって、肺の過剰炎症と重度の呼吸不全で特徴づけられる"急性呼吸窮迫症候群(ARDS)"という致死的病態を惹き起こしますが、その発症メカニズムは十分解明されていません。また現在使用されている抗インフルエンザ薬は、ARDSが惹き起こされたインフルエンザには効果がありません。さらに強毒型ウイルスによるパンデミックが発生した場合は、多数の死者が出て社会活動の崩壊の起こることが懸念されています。私達は、重症化を予測し、それに基づいた先制医療を行うことができれば、致死的病態に陥ることを阻止できるのではないかと考えています。ウイルスと宿主の相互作用は、動的で多階層的な生命現象です。そして、これらの相互作用の結果、宿主におけるウイルスの病原性の発現が決定されます。私達は、インフルエンザの重症病態を予測するために、これまでのインフルエンザ研究で既にデータが蓄積されているトランスクリプトミクスに加え、その上位あるいは下位に位置付けられるエピジェネティクスやメタボロミクス、さらにウイルス・宿主のインタラクトームなどの視点を取り入れたマルチスケールな解析を進めています。そして、これらのデータを基に、インフルエンザの重症病態をマルチスケールに数理モデリングする研究を進めています。これらの数理モデルと計算機シミュレーションを基に、多因子の重症化のバイオマーカーや治療標的を探索し、重症化や治療標的の予測を目指しています。
5. 肺貪食細胞由来NPYとその受容体の抗インフルエンザ作用
インフルエンザは、毎年冬季に流行する弱毒型のウイルスであっても、高齢者あるいはがん、肥満、糖尿病、喘息などの基礎疾患を有するヒトでは、生命に危険が及ぶほどに重症化して死に至ることがあります。しかしながらこれまでにインフルエンザが重症化するメカニズムや重症化を抑える有効な治療法は確立されていません。神経系と免疫系が相互に連関していることは知られていましたが、神経ペプチドがインフルエンザウイルス感染症の病態にどのように関わっているかは十分解明されていませんでした。神経ペプチドNPYは肥満、糖尿病、喘息などの病態に関わっており、NPYの受容体阻害薬は抗肥満薬として開発が進められています。しかしながら、NPYとその受容体がインフルエンザの重症化にどのように関わっているかは不明でした。一方、免疫応答など生体の恒常性の維持に必須のサイトカインのネガティブフィードバック因子であるSOCS3は強毒型のH5N1インフルエンザウイルス感染症やエボラ出血熱などの病態に関わっていることは報告されていましたが、神経ペプチドとの関わりは不明でした。
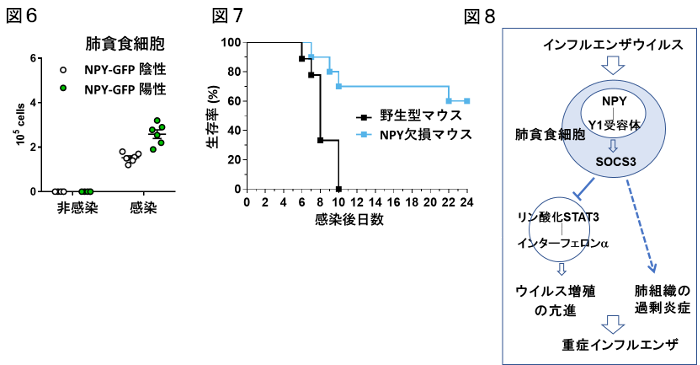
私達は、重症インフルエンザウイルス感染症に罹患したNPY遺伝子が活性化するとGFP蛍光を発現するマウスを使って、インフルエンザウイルスの感染に伴って肺の貪食細胞から神経ペプチドNPYが大量に産生されることを見出しました(図6)。またNPYとその受容体を貪食細胞で欠損させたマウスはインフルエンザの重症化が抑えられ、感染後の生存率が改善することがわかりました(図7)。そのメカニズムとして、ウイルス感染によってNPYとその受容体Y1R軸が活性化されると、サイトカインのネガティブフィードバック因子であるSOCS3の誘導を介して、ウイルス増殖の亢進と肺組織の過剰炎症が誘導され、インフルエンザが重症化することがわかりました(図8)。従って、貪食細胞におけるNPY-Y1R-SOCS3経路は重症インフルエンザの新しい治療標的となる可能性が考えられました。また、NPYはインフルエンザの重症化のバイオマーカーとして有用であると思われ、これを指標にインフルエンザの重症化が予測される患者に対して重症化を阻止するような予防医療の確立・先制医療の開発に繋がる可能性が示唆されました。これらの研究成果は、2018年11月『Nature Microbiology』に掲載されました。